Dan Benesh - Personal Site

Contact
University of California, Santa Barbara
Marine Science Institute
Santa Barbara 93106, USA
Email address: dbenesh82@gmail.com
Education and Employment
DFG Fellowship - 2013-2017
University of California, Santa Barbara
Research focus: Parasite life cycles in the food web
Postdoctoral researcher - 2007-2013
Max Planck Institute for Evolutionary Biology, Plön, Germany
Research focus: The evolution of complex parasite life cycles
M.S. and Ph.D - 2004-2007
University of Jyväskylä, Finland
Thesis topic: Larval life history strategies of an acanthocephalan parasite
B.S. - 2000-2004
University of Nebraska, Lincoln, U.S.A.
Major: Biological Sciences, Minors: Philosophy and Entomology
My CV



Research Interests
I am interested in the evolutionary ecology of parasites, mainly helminths (i.e. worms). My recent research has focused on the evolution of complex parasite life cycles and addresses two main questions: 1) why have complex life cycles evolved and 2) how do parasites successfully complete their life cycles?
1. Many parasites, including some of medical and veterinary importance, obligatorily use several hosts in their life cycle, i.e. they have a complex life cycle. At first glance, complex life cycles are puzzling, because a simple, one-host life cycle seems intuitively easier to complete. Parasites also must adapt to multiple host physiologies and immune systems. However, there are potential advantages for parasites with a complex life cycle, such as avoiding mortality from predation, achieving higher transmission rates, reaching a larger size at maturity, and increasing the probability to find a mate. Such benefits have rarely been shown in practice, though.
2. How should complex life cycle parasites divide their life among their multiple hosts? How much time should a worm spend growing and developing in one host before switching to the next host? And what are the costs and benefits that determine this? Most models addressing such questions are based in life history theory, and thus imply that parasite growth and mortality rates in consecutive hosts should shape the parasite strategy. A main prediction is that the size and age at which parasites are transmitted between hosts maximizes lifetime growth while minimizing mortality.
I use both cross-species comparative analyses and experiments to explore these questions. During my postdoc at the MPI in Plon, I used the tapeworm Schistocephalus solidus as a model system. It has three hosts in its life cycle (a copepod, a stickleback, and a piscivorous bird). Through experimental infections, I discovered that big worms in copepods are better at infecting sticklebacks (Benesh et al. 2012. Oikos), mainly because fast-growing worms also develop more rapidly (Benesh and Hafer 2012. Parasit. Vec.). The entire life cycle of S. solidus can be conveniently run in the laboratory, because the bird final host can be replaced with an in vitro system. The ability to breed worms allows us to perform studies that are impossible for most parasites, such as parentage and quantitative genetic studies. Some findings include: (i) high variation in paternity after indirect competition for fertilizations (Andreou and Benesh 2014. Parasitology), (ii) significant non-genetic parental effects on larval performance (Benesh 2013. J. Evol. Biol.), (iii) strong lifetime inbreeding depression and its potential to be purged through continued inbreeding (Benesh et al. 2014. Evolution).
The comparative approach has also yielded insights into Schistocephalus’s strategy, like why it refrains from growth in birds (because they undergo massive growth in fish, Benesh et al. 2013. Am. Nat.). Our broader cross-species comparisons have revealed some general patterns. For example, helminths that are transmitted along similar trophic links (e.g. copepod – fish or insect – terrestrial vertebrate) have frequently converged on similar life history strategies, i.e. they have evolved similar size and age at infectivity (Benesh et al. 2011. Evolution). And in nematodes, species that infect high trophic level definitive hosts tend to have longer life cycles, suggesting that infecting top predators favors the incorporation and/or maintenance of intermediate hosts in the life cycle (Benesh et al. 2014. Proc. R. Soc. B).

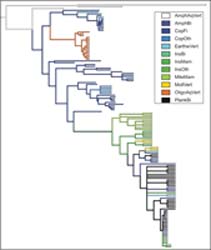
Recent Research
From 2014 to 2016, I worked on a DFG funded project to compile life cycle information for parasitic helminths. The resulting database summarizes decades of research and includes nearly 1000 species of thorny-headed worms, tapeworms, and roundworms (Benesh et al. 2017. Ecology). These data can be used to test hypotheses about life cycle evolution, host specificity, parasite life-history strategies, and the roles of parasites in food web. More simply, they document the incredible diversity in parasite strategies. For example, the figure below shows how body size changes over the life cycles for different species. In some hosts, parasites grow considerably whereas in others they just wait to be transmitted without growing at all. Some of the factors underlying this diversity can be explored in an interactive chart I built.

Publications
Contact me for pdfs or download them from my ResearchGate profile.
Check out my citation statistics here.
Benesh, D.P., Lafferty, K., and Kuris, A. 2017. A life cycle database for parasitic acanthocephalans, cestodes, and nematodes. Ecology. In Press. doi: 10.1002/ecy.1680
Benesh, D.P. 2016. Autonomy and integration in complex parasite life cycles. Parasitology 143: 1824-1846. https://doi.org/10.1017/S0031182016001311 (invited review)
Benesh, D.P. and Kalbe, M. 2016. Experimental parasite community ecology: intraspecific variation in a large tapeworm affects community assembly. Journal of Animal Ecology 85 (4): 1004–1013. doi: 10.1111/1365-2656.12527
Hafer, N. and Benesh, D.P. 2015. Does resource availability affect host manipulation? – an experimental test with Schistocephalus solidus. Parasitology Open 1: e3. doi: http://dx.doi.org/10.1017/pao.2015.3
Tuomainen, A., Valtonen, E.T., and Benesh, D.P. 2015. Sexual segregation of Echinorhynchus borealis (Acanthocephala) in the gut of burbot (Lota lota). Folia Parasitologica 62: 061. doi: 10.14411/fp.2015.061
Benesh, D.P., Aura, R.-L., Andersin, A.-B., and Valtonen, E.T. 2015. The occurrence of Echinorhynchus salmonis Müller, 1784 in benthic amphipods in the Baltic Sea. Folia Parasitologica 62: 052. doi: 10.14411/fp.2015.052
Aura, R.-L., Benesh, D.P., Palomäki, R., and Valtonen, E.T. 2015. The natural history of Echinorhynchus bothniensis Zdzitowiecki and Valtonen, 1987 (Acanthocephala) in a high Arctic lake. Folia Parasitologica 62: 051. doi: 10.14411/fp.2015.051
Benesh, D.P., Chubb, J.C., and Parker, G.A. 2014. The trophic vacuum and the evolution of complex life cycles in trophically-transmitted helminths. Proceedings of the Royal Society B. 281: 20141462. doi: 10.1098/rspb.2014.1462
Benesh, D.P., Weinreich, F., Kalbe, M., and Milinski, M. 2014. Lifetime inbreeding depression, purging, and mating system evolution in a simultaneous hermaphrodite tapeworm. Evolution 68 (6): 1762-1774. doi: 10.1111/evo.12388
Weinreich, F., Kalbe, M., and Benesh, D.P. 2014. Making the in vitro breeding of Schistocephalus solidus more flexible. Experimental Parasitology 139: 1-5. doi: 10.1016/j.exppara.2014.02.002
Andreou, D. and Benesh, D.P. 2014. Copulation order, density cues, and variance in fertilization success in a cestode. Parasitology 141 (7): 934-939. doi:10.1017/S0031182014000043
Benesh, D.P. 2013. Parental effects on the larval performance of a tapeworm in its copepod first host. Journal of Evolutionary Biology 26 (8): 1625-1633. doi: 10.1111/jeb.12165
Henrich, T., Benesh, D.P., and Kalbe, M. 2013. Hybridization between two cestode species and its consequences for intermediate host range. Parasites & Vectors. 6: 33. doi: 10.1186/1756-3305-6-33
Benesh, D.P., Chubb, J.C., and Parker, G.A. 2013. Complex life cycles: why refrain from growth before reproduction in the adult niche? The American Naturalist 181: 39-51. doi: 10.1086/668592
Weinreich, F., Benesh, D.P., and Milinski, M. 2013. Suppression of predation on the intermediate host by two trophically-transmitted parasites when uninfective. Parasitology 140: 129-135. doi: 10.1017/S0031182012001266
Benesh, D.P. and Hafer, N. 2012. Growth and ontogeny of the tapeworm Schistocephalus solidus in its copepod first host affects performance in its stickleback second intermediate host. Parasites & Vectors 5: 90. doi: 10.1186/1756-3305-5-90
Benesh, D.P., Weinreich, F., and Kalbe, M. 2012. The relationship between larval size and fitness in the tapeworm Schistocephalus solidus: bigger is better? Oikos 121: 1391-1399. doi: 10.1111/j.1600-0706.2011.19925.x
Benesh, D.P., Chubb, J.C., and Parker, G.A. 2011. Exploitation of the same trophic link favors convergence of larval life history strategies in complex life cycle helminths. Evolution 65: 2286-2299. doi: 10.1111/j.1558-5646.2011.01301.x
Benesh, D.P. 2011. Intensity-dependent host mortality: what can it tell us about larval growth strategies in complex life cycle helminths? Parasitology 138: 913-925. doi: 10.1017/S0031182011000370
Benesh, D.P. 2010. What are the evolutionary constraints on larval growth in a trophically-transmitted parasite? Oecologia 162: 599-608. doi: 10.1007/s00442-009-1507-6.
Benesh, D.P. 2010. Developmental inflexibility of larval tapeworms in response to resource variation. International Journal for Parasitology 40 (4): 487-497. doi: 10.1016/j.ijpara.2009.10.001
Benesh, D.P., Seppälä, O., and Valtonen, E.T. 2009. Acanthocephalan size and sex affect the modification of intermediate host coloration. Parasitology 136 (8): 847-854. doi: 10.1017/S0031182009006180
Hasu, T., Benesh, D.P., and Valtonen, E.T. 2009. Differences in parasite susceptibility and cost of resistance in naturally exposed and unexposed host populations. Journal of Evolutionary Biology 22 (4): 699-707. doi: TT10.1111/j.1420-9101.2009.01704.x
Benesh, D.P., Hasu, T., Seppälä, O., and Valtonen, E.T. 2009. Seasonal changes in host phenotype manipulation by an acanthocephalan: time to be transmitted? Parasitology 136 (2): 219-230. doi: 10.1017/S0031182008005271
Seppälä, O., Valtonen, E.T., and Benesh, D.P. 2008. Host manipulation by parasites in the world of dead-end predators: adaptation to enhance transmission? Proceedings of the Royal Society B 275: 1611-1615. doi: 10.1098/rspb.2008.0152
Benesh, D.P., Valtonen, E.T., and Seppälä, O. 2008. Multidimentionality and intra-individual variation in host manipulation by an acanthocephalan. Parasitology 135 (5): 617-626. doi: 10.1017/S0031182008004216
Benesh, D.P., Kitchen, J., Pulkkinen, K., Hakala, I., and Valtonen, E.T. 2008. The effect of Echinorhynchus borealis (Acanthocephala) infection on the anti-predator behavior of a benthic amphipod. Journal of Parasitology 94 (2): 542-545. doi: 10.1645/GE-1380.1
Benesh, D.P., Valtonen, E.T., and Jormalainen, V. 2007. Reduced survival associated with precopulatory mate guarding in male Asellus aquaticus (Isopoda). Annales Zoologici Fennici 44 (6): 425-434.
Benesh, D.P. and Valtonen, E.T. 2007. Effects of Acanthocephalus lucii (Acanthocephala) on intermediate host survival and growth: implications for exploitation strategies. Journal of Parasitology 93 (4). 735-741. doi: 10.1645/GE-1093R.1
Benesh, D.P. and Valtonen, E.T. 2007. Proximate factors affecting the larval life history of Acanthocephalus lucii (Acanthocephala). Journal of Parasitology 93 (4). 742-749. doi: 10.1645/GE-1094R.1
Benesh, D.P. and Valtonen, E.T. 2007. Sexual differences in larval life history traits of acanthocephalan cystacanths. International Journal for Parasitology 37 (2): 191-198. doi: 10.1016/j.ijpara.2006.10.002
Benesh, D.P., Hasu, T., Suomalainen, L.R., Valtonen, E.T., and Tiirola, M. 2006. Reliability of mtDNA in an acanthocephalan: the problem of pseudogenes. International Journal for Parasitology 36 (2): 247-254. doi: 10.1016/j.ijpara.2005.09.008
Benesh, D.P., Duclos, L.M., and Nickol, B.B. 2005. Behavioral response of amphipods harboring Corynosoma constrictum (Acanthocephala) to various components of light. Journal of Parasitology 91 (4): 731-736. doi: 10.1645/GE-440R.1